Происхождение эукариот

Бактерии и археи населяют всю поверхность планеты, толщу и дно океанов и несколько километров земной коры. Однако рекорды размеров и сложности принадлежат другому домену живых организмов — эукариотам. К ним относятся животные и растения, водоросли, грибы и разнообразные одноклеточные организмы — инфузории, амебы, жгутиконосцы и другие. Клетки эукариот отличаются от клеток бактерий и архей во многих отношениях (рис. 1). Они крупнее примерно в десять раз, содержат ядро, отделяющее геном и его транскрипцию от процессов цитоплазмы. В клетках эукариот имеются сложные системы мембранных цистерн и пузырьков — эндоплазматический ретикулум, аппарат Гольджи, лизосомы и другие вакуоли. Пузырьки управляемо отшнуровываются от мембран в одних местах и сливаются с ними в других. Белки цитоскелета — актин и тубулин — поддерживают форму клетки, а связанные с ними моторные белки — миозин, динеин, кинезин — перемещают органеллы внутри клетки и обеспечивают движение всей клетки. Если повредить мембрану бактериальной клетки, то все ее содержимое начнет вытекать наружу. Из поврежденной эукариотической клетки вытекают только вода и малые молекулы, а крупные белки и РНК остаются внутри, потому что они прикреплены к цитоскелету.
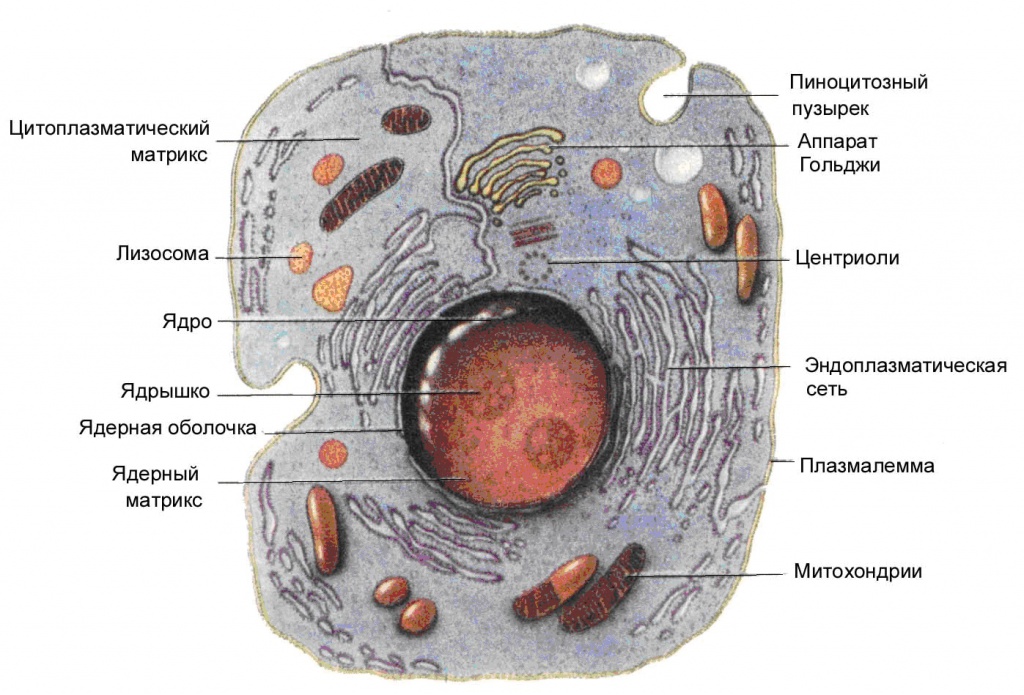
|
| 1. Эукариотическая клетка |
На уровне внутриклеточной молекулярной машинерии между бактериями и эукариотами также есть существенные отличия. Рибосомы эукариотической клетки крупнее, чем бактериальные, содержат больше белков и узнают матричные РНК по наличию кэпа — специальной модификации 5'-конца мРНК. Кроме кэпа эукариотические мРНК имеют полиадениловый хвост на 3'-конце, и, в отличие от бактериальных, каждая из них кодирует только один белок. Дыхательные цепи и мембранные АТФазы убраны с плазматической мембраны в специальные органеллы — митохондрии и пластиды. Геном эукариот состоит из нескольких линейных молекул ДНК — хромосом, намотанных для компактности на специальные белковые «катушки» — гистоны. Размер генома может достигать десятков миллиардов пар нуклеотидов, что на четыре порядка больше, чем у бактерий. Геном часто переполнен некодирующими последовательностями разных типов, из которых шире всего распространены так называемые сплайсеосомные интроны. При делении клетки цитоскелет разделяет реплицированные хромосомы по дочерним клеткам — эти сложные перемещения напоминают эпизод классического балета.
Наконец, в жизненном цикле многих эукариот есть половой процесс, собирающий гены двух родителей в одной клетке, и мейоз — специальное деление, при котором хромосомы, происходящие от разных родителей, обмениваются отдельными участками, после чего диплоидная клетка делится на четыре гаплоидных (с одиночным хромосомным набором). Благодаря мейозу образуются половые клетки.
Таким образом, эукариотическая клетка гораздо сложнее организована, чем бактериальная, и ее появление было таким же крупным эволюционным событием, как переход от РНК-мира к первым клеткам.
Первым шагом к пониманию происхождения эукариотической клетки стало обнаружение сходства между митохондриями и свободноживущими аэробными бактериями, а также между пластидами и цианобактериями. Первые работы в этом направлении появились еще в конце XIX века, а в современном виде теорию симбиогенеза сформулировала Линн Маргулис в 1970-х годах. К тому времени уже было обнаружено, что пластиды и митохондрии имеют собственную миниатюрную генетическую систему, включающую в себя кольцевую молекулу ДНК, а также особые рибосомы — мельче, чем в цитоплазме эукариот, и похожие на бактериальные. Органеллы никогда не образуются «с нуля», а только путем роста и деления существующих. Все это указывает на происхождение пластид и митохондрий от бактериальных симбионтов, когда-то поселившихся в цитоплазме эукариотической клетки.
Тогда же, в 1970-е, предпринимались попытки расширить теорию симбиогенеза и на другие системы эукариотической клетки. Несколько раз сообщалось об обнаружении собственной ДНК в центриолях — центрах организации тубулинового цитоскелета, в гидрогеносомах и пероксисомах, — специальных вакуолях, осуществляющих реакции с выделением водорода и утилизацию перекиси водорода. С применением более совершенных методов эти открытия пришлось «закрыть», но оказалось, что пероксисомы и гидрогеносомы почему-то получают новые липиды для мембран по одной молекуле через специальную систему транспортных белков, ту же, что доставляет липиды в митохондрии. В то же время обычные вакуоли получают новые липиды в виде целых мембранных пузырьков. Эта особенность пероксисом может означать, что они произошли от митохондрий путем крайнего упрощения и полной потери генома.
Гораздо сложнее было установить природу ядерно-цитоплазматического компонента (ЯЦК) эукариотической клетки — то есть «хозяина» митохондрий и пластид, или всей остальной части клетки, кроме них. Сегодня, когда прочитаны последовательности геномов более чем тысячи видов бактерий и архей, появилась возможность методами сравнительной геномики искать прокариотные гомологи каждого из тысяч эукариотических белков (см., например, А.Марков, А.Куликов, «Происхождение эвкариот: выводы из анализа белковых гомологий», в сборнике «Происхождение и эволюция биосферы», 2005, Новосибирск: ИК РАН). Краткие результаты такого поиска представлены на рисунках 2, 3. (Домены — относительно независимые части белковой молекулы. Новые белки могут возникать путем комбинирования старых доменов, и общие домены бывают в очень разных белках. Поскольку перестановки и новые комбинации доменов происходили много раз в эволюции эукариот, прослеживать эволюционную судьбу доменов проще, чем судьбу целых белков.)
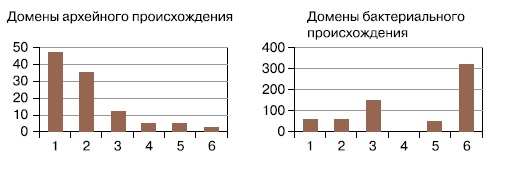
|
||
|
2. Вот как распределяются по функциональным группам «архейные» и «бактериальные» домены эукариотических белков: 1 — синтез белка, 2 — репликация, транскрипция, модификация и организация наклеиновых кислот, 3 — сигнальные и регуляторные белки, 4 — белки, связанные с образованием и функционированием мембранных пузырьков, 5 — транспортные и сортировочные белки, 6 — метаболизм (http://evolbiol.ru/dok_ibr2009.htm)
|
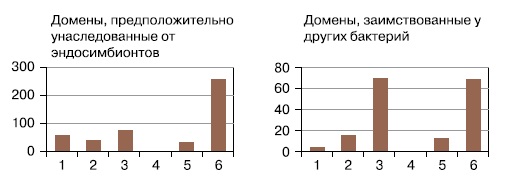
Хорошо видно, что доля архейных белков максимальна в обслуживании базовых генетических процессов эукариотических клеток — репликации, транскрипции, трансляции, репарации. Гены с этими функциями реже всего подвергаются горизонтальному переносу и, скорее всего, были унаследованы эукариотами напрямую. Гены бактериального происхождения, относящиеся к этим функциональным группам, в основном работают в митохондриях и пластидах и явно перенесены из их геномов. Это значит, что предок ЯЦК, скорее всего, был близок к археям.
Происхождение митохондрий благодаря сравнительной геномике удалось сильно уточнить. Их бактериальные предки относились к группе альфа-протеобактерий, причем в современном мире к ним ближе всего находятся пурпурные бактерии Rhodospirillum. Эти бактерии способны как к фотосинтезу, причем используют соединения серы для восстановления CO2, так и к аэробному дыханию на готовой органике, переключаясь между этими режимами метаболизма в зависимости от наличия кислорода и света. Большая часть их дыхательной цепи работает и в фотосинтезе, и в аэробном дыхании.
Некоторые эукариоты — паразитические простейшие Giardia, микроспоридии — не имеют митохондрий. Ранее считалось, что они никогда их и не имели, поскольку отделились от остальных в глубокой древности. Сегодня выясняется, что в их ядерном геноме есть гены альфа-протеобактериального происхождения, а значит, митохондрии у них были, но впоследствии исчезли.
Одна из малоизвестных функций митохондрий — сборка железосерных кластеров ферментов, в том числе работающих в цитоплазме (Tielens, A.G.M. et al, «Mitochondria as we don’t know them», «Trends in Biochemical Sciences», 2002, 27, 564—572, doi: 10.1016/S0968-0004(02)02193-X). Для этого требуется транспорт свернутых белковых глобул, содержащих железосерные кластеры, через митохондриальную мембрану. Это достаточно сложное и затратное приспособление, которое могло возникнуть, только если хозяин митохондрий сам не умел собирать железосерные кластеры.
Простейшее предположение о природе белков бактериального происхождения, работающих в цитоплазме эукариот, — что они все принадлежали предку митохондрий или пластид, — оказывается неверным. У эукариот насчитывается более 200 генных семейств бактериального происхождения, отсутствующих у альфа-протеобактерий и цианобактерий и, следовательно, приобретенных иными путями. Другие белки эукариот хотя имеют альфа-протеобактериальные гомологи, но ближе к белкам других групп бактерий. Например, все ферменты гликолиза эукариот наиболее сходны с белками бактерий-бродильщиков рода Clostridium.
Несколько белковых семейств эукариот, связанных со слиянием и отшнуровкой мембран, имеют архейное происхождение — например, адаптин N, синтаксин, а также белки, содержащие ароматический диаланиновый повтор, то есть те самые SNAP, за открытие которых вручили Нобелевскую премию 2013 года (см. статью в этом же номере). К сожалению, их функции в клетках архей неизвестны.
Помимо митохондрий для приспособления к кислороду нужны белки, защищающие цитоплазму, такие, как оксигеназы, пероксидазы, каталаза. Эукариоты получили все эти белки от бактерий, причем не родственных митохондриям. Этого и следовало ожидать — ведь чтобы получать выгоду от аэробного дыхания, надо сначала научиться защищаться от токсического действия кислорода, которое проявляется уже в очень малых концентрациях.
Мембраны эукариот, подобно бактериальным, состоят из сложных эфиров жирных кислот. Ферменты биосинтеза липидов соответственно могли быть получены только от бактерий. Отличительная черта мембран эукариотических клеток — вспомогательные липиды, стеролы, такие, как холестерин. Стеролы повышают текучесть мембран и поэтому необходимы для фагоцитоза и транспорта мембранных пузырьков. Следовательно, освоение биосинтеза стеролов должно было предшествовать появлению митохондрий. Предшественник всех стеролов — терпеновый углеводород сквален, который мы уже упоминали в главе, посвященной эволюции мембран. Синтез сквалена происходит у многих бактерий и архей, тогда как дальнейшие стадии синтеза стеролов характерны только для эукариот (рис. 4).
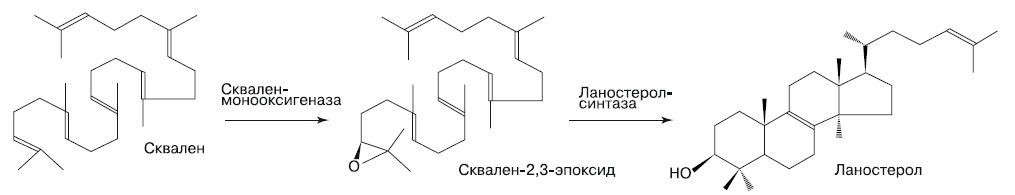
|
| 4. Биосинтез ланостерола — предшественника других стеролов (Lamb et al, 2007) |
На первом шаге сквален-монооксигеназа катализирует присоединение кислорода к концевой двойной связи сквалена, образуя сквален-2,3-эпоксид. Для этой реакции требуется молекулярный кислород, хотя и в очень малой концентрации. Затем ланостерол-синтаза катализирует восстановление эпоксида, при этом неустойчивый продукт восстановления — радикал сквалена — перестраивает двойные связи в замкнутые кольца, образуя ланостерол (у животных и грибов) или циклоартенол (у растений и миксомицетов).
Из-за крайней важности стеролов для эукариотической клетки происхождение ферментов их биосинтеза внимательно исследовалось. Сходные ферменты, синтезирующие те же стеролы, были обнаружены у некоторых бактерий, например у Methylococcus capsulatus из гамма-протеобактерий (Lamb et al,, «Molecular Biology and Evolution», 2007, 24, 8, 1714—1721, doi: 10.1093/molbev/msm090), Gemmata obscuriglobus из планктомицетов (Pearson et al, «Proceedings of the National Academy of Sciences USA», 2003, 100, 26, 15352—15357, doi: 10.1073/pnas.2536559100) и у ряда микобактерий, например Plesiocystis pacifica и Stigmatella aurantiaca. Однако подробное исследование показало, что все эти ферменты бактерии получили горизонтальным переносом от эукариот уже после расхождения линий растений и животных (Desmond, Gribaldo, «Genome Biology and Evolution», 2009, 1, 364—381, doi: 10.1093/gbe/evp036). Первый шаг биосинтеза стеролов — присоединение кислорода к сквалену — мог быть поначалу одним из способов защиты от молекулярного кислорода, и лишь потом нашлось применение полученному сквален-2,3-эпоксиду.
Разумеется, это не всё. Большая часть эукариотических генов бактериального происхождения, не принадлежавших предкам митохондрий и пластид, кодирует защитные, рецепторные, транспортные, сигнальные и регуляторные белки. Сюда относятся, например, белки теплового шока Hsp90, белки TerC и Ttg2, связанные с устойчивостью к тяжелым металлам, семейство Ras, передающее сигналы в протеинкиназных путях, и др. Что интересно, белки, которые у эукариот обеспечивают взаимодействия между компонентами клетки, у бактерий нужны для связей между разными клетками в сообществе. Например, Tim44 эукариот обеспечивает транспорт белков через внутреннюю мембрану митохондрий из цитоплазмы, а его бактериальные гомологи экспортируют белки из клеток во внешнюю среду. А значит, когда возникла эукариотическая клетка, под одной мембраной оказались уже отработанные системы связи клеток в сообществе.
Происхождение цитоскелетных белков прояснилось буквально в последние три года. Ранее были известны белки FtsZ, отдаленно похожие по последовательности на тубулин, — они широко распространены у бактерий и архей, могут полимеризоваться в пластины и трубки и необходимы для деления прокариотических клеток. В 2012 году были найдены артубулины — близкие гомологи тубулина — у двух видов архей рода Nitrosoarchaeum, принадлежащих к группе Thaumarchaeota. Их функция пока неизвестна (Yutin, Koonin. «Biology Direct», 2012, 7, 10, doi: 10.1186/1745-6150-7-10).
Аналогичным образом у бактерий и архей широко распространены отдаленные гомологи актина — белки MreB, полимеризующиеся в тонкие нити и участвующие в перетяжке делящейся бактериальной клетки. Другие дальние гомологи актина кодируются плазмидами и служат для их расхождения при делении клетки. И только в 2009 году у нескольких видов группы Crenarchaeota были обнаружены кренактины — близкие гомологи эукариотического актина (Ettema et al, «Molecular Microbiology», 2011, 80, 4, 1052—1106, doi: 10.1111/j.1365-2958.2011.07635.x). Кренактины образуют нити, которые придают клеткам палочковидную форму, часто с разветвлениями. По своей последовательности кренактины близки не только к актинам, но и белкам Arp2/Arp3 (Actin-related proteins), которые у эукариот образуют разветвления актиновых филаментов. В некоторых случаях совместная полимеризация актина и Arp2/Arp3 на плазматической мебране достаточна для отшнуровки фагоцитозного пузырька (Yutin et al, «Biology Direct» 2009, 4, 9, doi: 10.1186/1745-6150-4-9).
Наряду с артублинами у Thaumarchaeota обнаружились гомологи еще одной эукариотической системы — ESCRT-III (endosomal sorting complex required for transport). Эта группа белков связана с мембранами комплекса Гольджи, эндоплазматического ретикулума и вакуолей и участвует в сортировке содержимого мембранных пузырьков. В клетках архей белки этого комплекса необходимы для деления клеток, заменяя отсутствующий FtsZ (Makarova et al, «Nature Reviews Microbiology», 2010, 8, 731—741, doi: 10.1038/nrmicro2406).
В геноме Caldiarchaeum subterranum, прочитанном в 2010 году, были обнаружены гомологи эукариотической системы убиквитиновой модификации белков. Эта система пришивает маленький белок убиквитин к различным клеточным белкам, что служит обычно меткой для их уничтожения в протеосоме, а также влияет на другие процессы, например перестройку хроматина. Кроме собственно убиквитина, в нее в минимальном варианте входят три фермента — Е1, Е2 и Е3, — катализирующие разные стадии его присоединения. У бактерий известны отдаленные гомологи Е1 и Е2, но их функции не связаны с убиквитином, они катализируют включение серы при синтезе витамина В1 и молибденовых кластеров ферментов. Протеосомы были известны ранее у некоторых бактерий (Mycobacterium tuberculosum) и архей (Haloferax volcanii), но убиквитиновой системы у этих видов нет. Протеосомы есть и у Caldiarchaeum subterranum, они похожи на эукариотические также по наличию белка RPN11, который опознает убиквитиновые метки на белках (Nunoura et al, «Nucleic Acids Research», 2011, 39, 3204–3223, doi: 10.1093/nar/gkq1228).
Еще один обязательный компонент эукариотической клетки, гистоновые белки, тоже унаследован от архей. ДНК эукариот намотана на нуклеосомы — комплексы из восьми гистоновых белков, по две молекулы гистонов H2A, H2B, H3 и H4; гистон H1 соединяет соседние нуклеосомы в цепочку. У некоторых архей есть гены гистонов, более всего сходные с эукариотическими H3 и H4, и образуются маленькие нуклеосомы из четырех белковых молекул, которые тоже служат для компактной укладки ДНК (Pereira et al, «Proceedings of the National Academy of Sciences USA», 1997, 94, 23, 12633—12637).
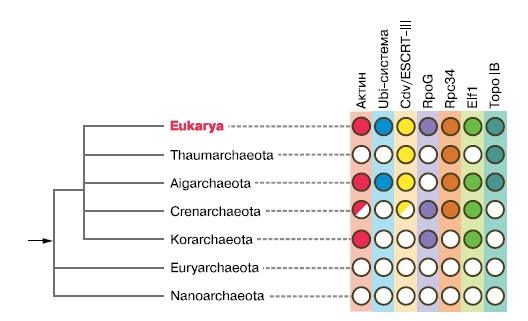
Распределение гомологов эукариотических систем среди различных архей позволило уточнить положение архейного предка эукариот. Современная система архей состоит из шести типов: Crenarchaeota, Euryarcheota, Nanoarchaeota, Thaumatoarchaeota, Aigarchaeota, Korarchaeota. Большинство известных видов относится к первым двум типам. Crenarchaeota — в основном обитатели горячих источников, чаще кислых, среди них есть как автотрофные хемосинтетики, так и гетеротрофы, осуществляющие сульфатное дыхание. К Euryarchaeota принадлежит большинство метаногенов, они широко распространены в почвах, болотах и морских осадках и предпочитают умеренные температуры, а также фотосинтезирующие обитатели пересоленых лагун, такие, как Halobacterium halobium. Автотрофные археи Thaumatoarchaeota, окисляющие аммиак и широко распространены в почвах К Aigarchaeota относится единственный подробно изученый вид, выделенный из горячих кислых вод золотого рудника, сильно отличающийся от Euryarcheota по молекулярным прихнакам. Korarchaeota — тоже малочисленная группа гетеротрофных архей из морских горячих источников, с тонкими длинными палочковидными клетками. Nanoarchaeota — очень мелкие шаровидные археи, живущие в симбиозе с другими археями в горячих источниках. Молекулярно-филогенетические методы показывают, что предкок эукариот был ближе к четырем типам группы ТАСК (Thaumatoarchaeota, Aigarchaeota, Crenarchaeota, Korarchaeota), а два других типа архей — Euryarchaeota и Nanoarchaeota — отделились раньше (Guy et al, «Trends in Microbiology» 2011. 19, 12, 580—587, doi: 10.1016/j.tim.2011.09.002).
|
|
|
Таким образом, мы видим, что ядерно-цитоплазматический компонент эукариотической клетки сочетает в себе черты архей и различных неродственных групп бактерий. От архей унаследованы центральные информационные системы клетки, зачатки цитоскелета, зачатки систем управления мембранами и убиквитиновая система модификации белков. От бактерий произошли ферменты метаболизма сахаров, липидов и отчасти — стеролов, системы защиты от кислорода и всевозможные сигнальные и регуляторные белки. Очевидно, что приобретение митохондрий было далеко не первым шагом на пути происхождения эукариот: чтобы такой симбиоз стал механически возможен, необходимы механизмы фагоцитоза. А чтобы он был еще и полезен, надо уже иметь ферменты гликолиза, защиту от кислорода, системы транспорта веществ и передачи сигналов между симбионтом и хозяином.
Как мы можем описать появление эукариот, исходя из всего сказанного выше? Во-первых, симбиогенез, многочисленные переносы генов от разных бактерий и большая роль сигнально-регуляторных генов означают, что процесс происходил в сложном сообществе прокариот. Во-вторых, архейное происхождение базовых генетических систем эукариот свидетельствует о том, что у истоков процесса стояли археи. Из разнообразных экологических групп современных архей только одна — метаногены — живет в сложных сообществах, в тесном взаимодействии с бактериями. Остальные археи, как правило, приспособлены к экстремальным местообитаниям, где образуют крайне бедные сообщества с малой ролью межвидовых взаимодействий. Отсутствие всяких следов метаногенеза у эукариот означает, что этот тип обмена веществ в какой-то момент эукариогенеза стал непригоден и был заменен другими. В качестве причины напрашивается появление кислородного фотосинтеза. Ферменты метаногенеза используют ионы двухвалентного никеля, которые очень уязвимы для кислорода.
Итак, скорее всего, эукариогенез начался с метаногенной археи, обитавшей в нижних слоях бактериального мата в симбиозе с ацетогенными бродильщиками, выделяющими водород. Появление кислородного фотосинтеза в верхнем слое того же мата вызвало крупный экологический кризис. Многие обитатели мата вымерли, другим же удалось создать свои или приобрести горизонтальным переносом чужие системы защиты от кислорода. Это произошло не мгновенно, процесс распространялся с поверхности в глубокие слои мата, и к моменту появления серьезных проблем у метаногенов их соседи сверху уже имели адаптации к кислороду. Многие прокариоты в состоянии стресса начинают активно поглощать ДНК из внешней среды, и таким способом наша архея приобрела гены, необходимые для защиты от кислорода и новый метаболизм, скорее всего, молочнокислое брожение. Сквален-монооксигеназа, фермент синтеза стеролов, могла исходно служить для защиты от кислорода. Отсутствие клеточной стенки, актиновый цитоскелет для поддержания вытянутой и ветвистой формы клеток, стеролы и освобождение мембраны от энергетических функций позволили архее перейти к фагоцитозу и успешно конкурировать с соседями-бактериями. Такой способ питания усилил поток чужеродных генов в цитоплазму клетки, что, видимо, потребовало специализации РНК-полимераз — появления трех из одной предковой.
Тем временем в среднем слое мата, среди пурпурных фотосинтезирующих бактерий, было отработано аэробное дыхание, ценным субстратом для которого стали органические кислоты, выделяемые бродильщиками нижних слоев. Эти аэробы стали оптимальными партнерами для симбиоза с фагоцитирующими бродильщиками. Вероятно, сначала взаимодействие между ними было отработано на стадии простого соседства в мате, а затем продолжилось с переходом к эндосимбиозу.
Поток чужеродных генов неизбежно должен был заражать фаготрофную клетку новыми видами вирусов и мобильных элементов. Их давление могло вызвать ответ в виде образования ядра для защиты генетического материала. Разделение клетки на компартменты, один из которых специализируется на хранении ДНК, известно и у бактерий группы Planctomycetes (Fuerst, Sagulenko, «Frontiers in Microbiology», 2012, 3, 167, doi: 10.3389/fmicb.2012.00167). К сожалению, чтение и аннотация их геномов пока не закончены.
У бактерий и архей синтез белков, закодированных в матричной РНК, начинается еще до того, как эта мРНК будет полностью синтезирована. Поэтому интроны встречаются у бактерий и архей в небольшом количестве, и только в генах рибосомных и транспортных РНК. В этом случае после окончания синтеза РНК у интрона есть время, чтобы вырезать себя из нее. При попадании интронов в белок-кодирующие гены бактерий вырезание интрона из мРНК будет осложнено — «голова» интрона окажется в рибосоме еще до того, как «хвост» будет достроен, интрон не сможет вырезаться из мРНК, и синтезированный с нее белок будет дефектным.
Появление ядра, разделившего трансляцию и транскрипцию, сняло эти ограничения, и началось бурное размножение интронов в белок-кодирующих генах предка эукариот. Мутации интронов часто нарушали их способность к самовырезанию и приводили к появлению большого количества дефектных белков. Это вызвало давление отбора на появление системы контроля качества мРНК (nonsence-mediated decay) и специализацию части интронов на вырезании других, поврежденных интронов. Так возникли малые ядерные РНК, составляющие основу сплайсосомы.
Новые мобильные элементы резко активизировали рекомбинацию, что, возможно, привело к распаду кольцевой ДНК на линейные хромосомы. Их концы при репликации могли сначала защищать мобильные элементы, размножающиеся параллельно делению клетки, подобно тому, как это сейчас происходит у ряда насекомых. Затем на основе обратной транскриптазы ретротранспозонов возникла теломераза.
После установления симбиоза с митохондриями протоэукариоты стали высокоэффективными хищниками: аэробное дыхание повышает эффективность использования пищи почти в 20 раз по сравнению с гликолизом. Они смогли выйти на поверхность мата и начать фагоцитировать цианобактерий. Так часть эукариот приобрела второй тип эндосимбионтов — хлоропласты. Они стали водорослями и ушли из бактериального мата в толщу океана, другие же остались гетеротрофами и продолжили питаться бактериями. В дальнейшем среди эукариот появились крупные хищные формы, такие, как инфузории, приспособившиеся к питанию другими эукариотами, в первую очередь водорослями. Они вместе со своей добычей составили новую экосистему открытого моря, независимую от занявших мелководья бактериальных матов. Наличие цитоскелета и сложной системы регуляции генов позволило эукариотным клеткам объединиться в крупные многоклеточные организмы. За этим последовали радикальное усложнение биосферы («кембрийский взрыв») и ускорение эволюции, и с тех пор облик Земли определяют многоклеточные растения и животные.
Элементы жизни: почему не кремний и не фтор
Фотохимия и «черные курильщики»
Пути восстановления углекислого газа
Происхождение белкового синтеза и генетического кода
Мир вирусов, последний общий предок и происхождение ДНК
Происхождение мембран и мембранной биоэнергетики
Закат «цинкового мира», прокариотная биосфера и происхождение фотосинтеза
Происхождение эукариот