Роль вирусов в происхождении клеточного ядра

В прошлом номере мы рассмотрели происхождение эукариот путем симбиоза между археей и бактерией в условиях перехода бактериального сообщества к аэробному обмену. Однако этот сценарий не объясняет всех особенностей эукариотических клеток. Непонятно, как переход археи к фагоцитозу и обзаведение симбионтами могли способствовать появлению, например, полового размножения и мейоза. Не очень понятно происхождение линейных хромосом. И совершенно несуразной выглядит такая деталь эукариотической системы трансляции, как кэпы матричных РНК. Кэп — это остаток 7-метилгуанина, пришитый к 5'-концу мРНК специальным 5'-5'-трифосфатным мостиком (рис. 1). Он требуется для начала трансляции на эукариотической рибосоме — без него рибосома не может связаться с мРНК.
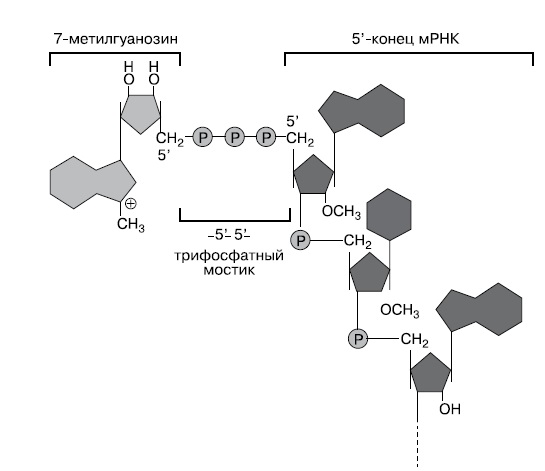
|
| 1. Структура 5' кэпа мРНК (wikipedia.org) |
Для узнавания кэпа служит специальный белок — фактор инициации eIF4E, не имеющий гомологов у прокариот. Важно, что из-за наличия кэпа и eIF4E эукариоты не имеют оперонов — групп генов, которые транскрибируются в одну длинную мРНК, кодирующую несколько белков последовательно, один за другим. Опероны позволяют бактериям и археям экономить на регуляции активности генов: достаточно одного регуляторного участка в начале оперона, чтобы одновременно управлять активностью всех его генов. (Обычно белки, которые кодируются генами одного оперона, функционально связаны, а значит, требуются в равных количествах.) Переход к кэпированным мРНК у протоэукариота потребовал бы массированной перестройки генома, распада всех оперонов и появления тысяч новых промоторов для индивидуализирующихся генов. Сложно представить себе причину, по которой это было бы адаптивно для клетки, и еще сложнее найти причину, по которой система пришивания и узнавания кэпа вообще возникла.
Эти особенности эукариот находят объяснение, если допустить, что в их появлении участвовали, помимо архейного и бактериального партнеров, еще и вирусы.
Вирусное происхождение отдельных компонентов эукариотической клетки не вызывает сомнения. Так, фермент теломераза происходит от обратной транскриптазы ретровирусов, а часть генетического аппарата митохондрий — ДНК-полимераза, РНК-полимераза и праймаза — от Т-четного бактериофага, встроенного в геном бактериального предка митохондрий. Однако есть и более радикальное мнение о роли вирусов в появлении эукариотической клетки — вирусная теория происхождения ядра.
Эту теорию в современном виде выдвинули вирусологи Филипп Джон Белл и Масахара Такемура (Bell, «Journal of Molecular Evolution», 2001, 53, 3, 251—256, doi: 10.1007/s002390010215; Takemura, «Journal of Molecular Evolution», 2001, 52, 5, 419—425, doi: 10.1007/s002390010171; Bell, «Annals of the New York Academy of Sciences», 2009, 1178, 91-105, doi: 10.1111/j.1749-6632.2009.04994.x). Они утверждают, что ядро эукариотической клетки происходит от крупного ДНК-вируса, заразившего древнюю архею, перешедшего в лизогенное состояние и постепенно взявшего клетку хозяина под полный контроль. (Лизогения — сосуществование бактериальной клетки с умеренным фагом, в отличие от лизиса — убийства фагом бактерии.) На протяжении 2000-х годов другие ученые практически не рассматривали эту теорию всерьез, но в последнее время она стала набирать популярность — так, к ее развитию подключился крупнейший французский вирусолог Патрик Фортер.
К крупным ДНК-вирусам (LNCDV, Large Nucleo-Cytoplasmic DNA Viruses) относятся, например, поксвирусы (из них широко известен возбудитель оспы) и мимивирусы (недавно открытая группа вирусов-паразитов амеб и морских водорослей). Они имеют крупный и сложный вирион, покрытый несколькими мембранами. Самая внешняя мембрана сливается с плазматической мембраной клетки при заражении, и в цитоплазму попадает вирион, окруженный двумя внутренними мембранами. Эти мембраны местами сливаются между собой, образуя некое подобие ядерных пор, они выстланы изнутри белками капсида, аналогичными белкам ядерной ламины. В такой форме вирус долго существует в цитоплазме, в нем происходят транскрипция, кэпирование и полиаденилирование РНК, экспорт зрелых РНК в цитоплазму через поры вириона. Активные вирионы вируса оспы раньше называли «мини-ядрами», а вирион мимивируса по размеру почти не уступает ядру клетки-хозяина.
Сходство LNCDV с ядром эукариот наблюдается по многим признакам:
— генетический материал ограничен двухслойной липидной оболочкой с белковым каркасом;
— транскрипция и трансляция разделены в пространстве (транскрипция идет внутри вириона, трансляция — в цитоплазме клетки-хозяина);
— мРНК активно экспортируется через поры в мембранной оболочке;
— геном состоит из линейных молекул ДНК с тандемными повторами на концах;
— расхождение дочерних молекул ДНК при делении может сопровождаться исчезновением оболочки.
Геном таких вирусов представлен линейной двухцепочечной ДНК длиной до 200 тысяч пар нуклеотидов у поксвирусов и более миллиона — у мимивирусов, что сравнимо с размером самых малых бактериальных геномов. Открытые в 2013 году пандоравирусы имеют геном размером до 2,5 миллионов пар нуклеотидов, что соответствует геному средней свободноживущей бактерии. Для репликации ДНК эти вирусы разбирают вирион, а по мере накопления вирусной ДНК в цитоплазме клетки собираются новые вирионы, которые окружаются впячиваниями эндоплазматического ретикулума и плазматической мембраны. Кэпирование, полиаденилирование, а часто и синтез дезоксирибонуклеотидов и восстановление рибозы при размножении этих вирусов осуществляют их собственные ферменты, без участия клеточных белков.
Проведенный Беллом филогенетический анализ гуанилилтрансфераз — ферментов пришивания кэпа — показывает, что ферменты современных вирусов образуют на дереве единую самостоятельную ветвь, а ферменты эукариот — другую. Перемешивания между ними нет, это означает, что не было многократных переносов гена между вирусами и их хозяевами.
Чтобы понять, какой из узлов дерева гуанилилтрансфераз самый древний и соответствует предковому ферменту, к ним добавлены родственные ферменты — АТФ-зависимые ДНК-лигазы. Это более древние белки, предковые по отношению к гуанилилтрансферазам, соответственно ближайший к ним узел гуанилилтрансферазной части дерева укажет нам первую гуанилилтрансферазу в истории. Оказывается, она принадлежала вирусам — одна ветвь ее потомков содержит ферменты поксвирусов, другая — фермент вируса ASF (африканской чумы свиней — African swine fever). Гуанилилтрансферазы эукариот происходят от фермента вирусной линии, давшей начало вирусу ASF. Это доказывает, что кэпирование мРНК, как и другие инновации в генетических системах, появилось сначала среди вирусов и лишь затем было заимствовано эукариотами (рис. 2).
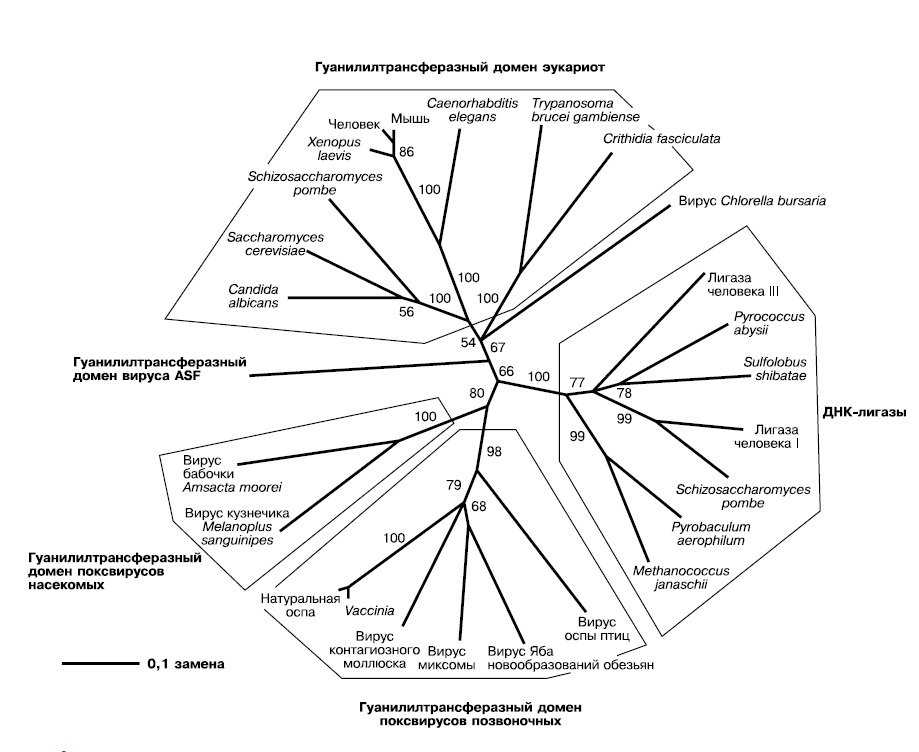
|
| 2. Филогенетическое дерево гуанилилтрансфераз (Bell, 2001) |
Для вирусов, в отличие от клеток, есть очевидная выгода в создании такой системы. Это средство перехвата управления рибосомами хозяина: белок — предок eIF4E присоединяется к рибосомам, после чего они узнают только кэпированные вирусные мРНК, останавливая синтез белков клетки. У мимивируса обнаружен собственный белок-аналог eIF4E, роль его пока непонятна. Аналогичным образом полиаденилирование РНК продлевает их существование в цитоплазме и тоже могло быть сначала адаптацией вируса. Возможно, вирусный фермент когда-то разрушал хозяйские матричные РНК, не имеющие поли-А хвоста.
Такемура обратил внимание на другой ключевой фермент эукариот, ДНК-полимеразу альфа. Как мы помним из предыдущих номеров, главная ДНК-полимераза эукариот, полимераза дельта, которая синтезирует основную часть новой ДНК, унаследована от архей. Полимераза альфа строит первые несколько десятков нуклеотидов новой цепи после РНК-праймера, после чего уступает место полимеразе дельта. В пробирке возможен синтез новой ДНК с помощью одной только полимеразы дельта, но в клетках эукариот этого не происходит. Полимераза альфа, судя по филогенетическим деревьям, получена предком эукариот от вирусов группы LNCDV, родственные, но менее сходные полимеразы встречаются у хвостатых бактериофагов.
Наконец, Патрик Фортер связал с вирусами происхождение огромного количества эукариотических белков, не имеющих гомологов среди бактерий и архей (Forterre, «Research in Microbiology», 2011, 162, 1, 77—91, doi: 10.1016/j.resmic.2010.10.005). По последним оценкам, общий предок современных линий эукариот уже имел более 1400 таких белков. Среди них преобладают вспомогательные белки цитоскелета и ДНК-связывающие белки, в структуре которых присутствуют многочисленные короткие повторы. Вирусы, особенно крупные, содержат много генов, не похожих ни на какие гены клеточных организмов, а часто — и на гены других вирусов. Кодируемые ими белки тоже часто имеют множественные короткие повторы. Низкая точность вирусных полимераз, интенсивная рекомбинация и гонка вооружений с хозяевами приводят к очень быстрой эволюции вирусных белков по сравнению с клеточными, поэтому вирусы — обильный источник принципиально новых белков.
По предложенному Беллом сценарию, вирус, ставший предком ядра, паразитировал на метаногенной архее с клетками разветвленной формы, не имевшей клеточной стенки (Bell, «Journal of Theoretical Biology», 2006, 243, 1, 54—63, doi: 10.1016/j.jtbi.2006.05.015). Такие археи есть и сейчас, например, Methanoplasma elizabethii; разветвленная форма позволяет им плотнее контактировать с выделяющими водород бактериями-ацетогенами. Как и у многих современных LNCDV, зрелые частицы этого вируса были покрыты тремя мембранами, внешняя из которых сливалась с мембраной заражаемой клетки, а две внутренние имели поры, пропускавшие РНК и другие молекулы из вируса в цитоплазму клетки и обратно. Такие мембранные оболочки современных вирусов строятся из цистерн эндоплазматического ретикулума клетки-хозяина, а у археи, не имевшей внутренних мембран, для их построения использовалась внешняя мембрана клетки. От мембраны клетки-хозяина отшнуровывались круглые пузырьки, которые затем складывались в двухслойное полушарие, окружали реплицированную вирусную ДНК и выходили из клетки, покрываясь при этом третьей мембраной (рис. 3). Подобным образом одеваются мембранами частицы бактериофага PRD1, и в его случае все манипуляции с мембранами производят только вирусные белки.
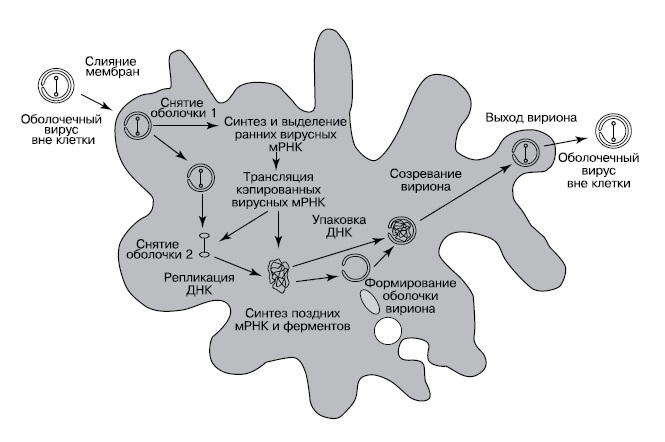
|
| 3. Предполагаемый цикл развития вируса — предка ядра (Bell, 2006) |
Ключевым моментом превращения вируса в ядро должен был стать, как мы уже упоминали, переход такого вируса в лизогенное состояние. Многие вирусы способны переключаться между литическим жизненным циклом (быстрое размножение с гибелью клеток хозяина) и лизогенным существованием — скрытой инфекцией, практически не влияющей на жизнеспособность хозяина. Лизогенное состояние достигается двумя способами: можно интегрировать свой геном в геном хозяина и пользоваться хозяйскими системами репликации или же, переключив активность вирусных генов, постоянно существовать в цитоплазме. В этом случае вирус временно превращается в плазмиду — автономную молекулу ДНК. Он тоже может пользоваться хозяйской системой репликации, но ему нужны собственные механизмы контроля количества копий. Если он будет отставать в репликации от генома хозяина, то деления хозяина будут часто порождать незараженные клетки, а если репликация вируса будет слишком активной, то он будет снижать жизнеспособность хозяина.
Большинство лизогенных бактериофагов и крупные плазмиды поддерживают свое число копий минимальным и используют специальную систему разделения копий по дочерним клеткам, похожей на ту, которая разделяет хромосомы в клетках современных эукариот. Эта система обычно включает в себя белок, обратимо полимеризующийся в нити, центромерные участки ДНК (то есть участки, к которым должны прикрепляться нити веретена деления и в которых соединяются сестринские хромосомы до того, как разойдутся по разным клеткам) и второй белок, связывающий центромеры с нитями первого белка. Запуск полимеризации нитей растаскивает две копии плазмиды или вирусной ДНК по двум концам удлиненной делящейся клетки. У некоторых плазмид, например R1, белок нитей похож на актин, у других встречаются гомологи тубулина. Плазмиды, не способные образовывать вирионы, имеют другой путь горизонтальной передачи в незараженные клетки — конъюгацию, образование специальных контактов между бактериальными клетками, по которым передается плазмидная ДНК.
И конъюгативные плазмиды, и лизогенные вирусы обычно способны отличить уже зараженные их собратьями клетки от «чистых», чтобы предотвратить бесполезное повторное заражение. Такое распознавание работает только в пределах близких видов вирусов, поэтому клетка может быть заражена одновременно несколькими разными лизогенными вирусами. Однако стабильная лизогенная инфекция несколькими вирусами требует, чтобы они использовали разные, несовместимые системы разделения копий по дочерним клеткам, иначе первое же деление клетки разносит разные лизогены по разным клеточным линиям.
Лизогенные вирусы, такие, как N15, и плазмиды, как R1, уже имеют цикл размножения, напоминающий митоз: они реплицируются однократно, после чего две копии остаются связанными в центромерном регионе. Дальнейшая репликация блокируется до тех пор, пока полимеризация нитей не растащит две копии далеко друг от друга. Гигантский вирус — предок ядра, в отличие от N15 и R1, имел оболочку вириона. Поэтому ему приходилось разбирать оболочку на время репликации и деления и восстанавливать ее для транскрипции генов, что еще ближе к митозу.
Следующим шагом от вируса к ядру стала потеря литического пути размножения. Вирус оказался «в одной лодке» с хозяином, и теперь давление отбора, во-первых, требовало минимизации вреда хозяину, во-вторых, создания механизма конъюгации для заражения новых хозяев. Второе было очень просто, так как вирус уже обладал белками, вызывающими слияние мембран. Достаточно было перенести их с оболочки вириона на плазматическую мембрану, чтобы, во-первых, получить возможность слияния клеток зараженного хозяина с незараженным и, во-вторых, улучшить способности хозяина к фагоцитозу за счет слияния двух его собственных псевдоподий вокруг добычи. На этой стадии использование кэпов и eIF4E для подавления трансляции генов хозяина стало опасным, и многие жизненно важные гены хозяина были перенесены в геном вируса под его управление. Современные мимивирусы содержат сотни генов, недавно перенесенных из генома хозяина, так что сценарий не выглядит нереальным. Затем кэпы пригодились для того, чтобы ограничить и поставить под контроль транскрипцию огромного количества генов, проникавших в цитоплазму из перевариваемых клеток с переходом к фагоцитозу. Поглощенные чужеродные ДНК могли транскрибироваться в цитоплазме, но получаемые при этом мРНК не имели кэпов и не узнавались рибосомами протоэукариотной клетки, оснащенными белком eIF4E. Необходимые митохондриальные гены переносились в ядро под контроль его систем регуляции. В итоге кольцевой геном архейного предка исчез совсем, а митохондриальные геномы сократились до считанных десятков генов.
Происхождение мейоза и полового цикла требует взаимодействия между разными штаммами такого лизогенного вируса. Два штамма, достаточно разных, чтобы опознавать друг друга как «чужих», но еще с одинаковыми системами сегрегации, могли вызывать слияние мембран своих клеток с последующей репликацией обоих вирусных геномов. Идентичность центромер и центромер-связывающих белков приводила к тому, что все четыре генома оказывались склеенными вместе по центромерам. Рекомбинационные белки, необходимые для достройки теломер вируса, могли в этот момент вызывать рекомбинации между геномами разных штаммов, аналогичные кроссинговеру. Деление хозяйской клетки приводило к разделению вирусных геномов по два в каждую дочернюю клетку, они оставались связанными по центромере и не реплицировались. Второе деление хозяина окончательно разделяло вирусные геномы и разрешало их репликацию. Естественный отбор мог закрепить такое поведение, потому что рекомбинация между двумя вирусными геномами в одной клетке позволяет им избавиться от вредных мутаций. Нечто подобное показано для вирусов с ультрафиолетовыми повреждениями после дезинфекции: при заражении одной клетки несколькими поврежденными вирусами из их геномов собирается один работоспособный и происходит успешное размножение (Barry, «Virology», 1961, 14, 4, 398—405, doi: 10.1016/0042-6822(61)90330-0).
Итак, вирусная теория происхождения ядра лучше других объясняет такие особенности эукариот, как кэпирование и полиаденилирование мРНК, отсутствие оперонов, митоз, мейоз и половые процессы. Она объясняет также происхождение систем слияния мембран и отшнуровки мембранных пузырьков. Серьезными доказательствами в ее пользу могут стать:
— обнаружение среди вирусов архей близких родственников LNCDV-вирусов;
— обнаружение у архейных вирусов системы кэпов;
— находка вирусных гомологов белков мейоза.
К сожалению, архейные вирусы пока остаются практически terra incognita.
Участие вируса в симбиозе, давшем начало эукариотам, снимает многие проблемы взаимодействия архейного и бактериального геномов в одной клетке и защиты от попадания чужеродной ДНК при фагоцитозе. Вирусы, особенно LNCDV, обладают изощренными механизмами для манипуляции чужими генетическими системами. Благодаря транскрипции внутри вирусной частицы, наличию кэпов и полиаденилированию матричных РНК вирус мог поддерживать функционирование зараженной клетки, несмотря на мощный поток чужих генов из фагоцитированных бактерий.
Используя вирусное наследие, эукариоты смогли подчинить сначала альфа-протеобактерий, ставших митохондриями, затем цианобактерий, ставших хлоропластами, а потом множество других бактерий. Всякий раз, когда эукариотам требовался какой-нибудь экзотический биохимический путь, они брали на работу бактерий, уже владеющих им. Так, десятки групп глубоководных животных независимо приручили хемосинтетические бактерии, окисляющие сероводород или метан. Азотфиксация, разложение целлюлозы, синтез витаминов и многое другое эукариоты делают благодаря симбиотическим бактериям. Более того, эукариоты способны вступать в эндосимбиоз с другими эукариотами. Гигантская группа водорослей, в том числе бурые, золотистые, диатомовые, криптофитовые, гаптофитовые и динофлагелляты, произошла от хищного жгутиконосца, приручившего одноклеточную красную водоросль. Следы этого симбиоза видны в четырехмембранных хлоропластах большинства представителей этих групп, а также в нуклеоморфе — маленьком остатке ядра красной водоросли между второй и третьей мембраной хлоропласта криптофитовых водорослей.
Вершиной же манипулятивных талантов вируса, ставшего ядром, можно считать клетку динофлагелляты Kryptoperidinium. Эта одноклеточная водоросль происходит от динофлагеллят, имевших хлоропласт, который был потомком красной водоросли, но затем перешедших к хищному образу жизни. Старый хлоропласт остался у них в качестве маленького фоторецептора. Потом эти хищные жгутиконосцы вступили в симбиоз с диатомовой водорослью, которая сохранила ядро и значительную часть генома. В клетке Kryptoperidinium под управлением ядра находятся еще пять геномов: митохондриальный, старого хлоропласта, ядро симбионта, митохондриальный геном симбионта и хлоропластный геном симбионта (рис. 4, Figueroa и др, «Protist», 2009, 160, 2, 285-300, doi: 10.1016/j.protis.2008.12.003 ). Деление ядер хозяина и симбионта строго синхронизировано. Более того, при половом размножении происходят мейоз и слияние как главных ядер половых клеток, так и ядер симбионтов. Наконец, в этом симбиозе может быть и четвертый уровень — иногда Kryptoperidinium сам становится симбионтом, поселяясь в клетках медузы Cassiopea!
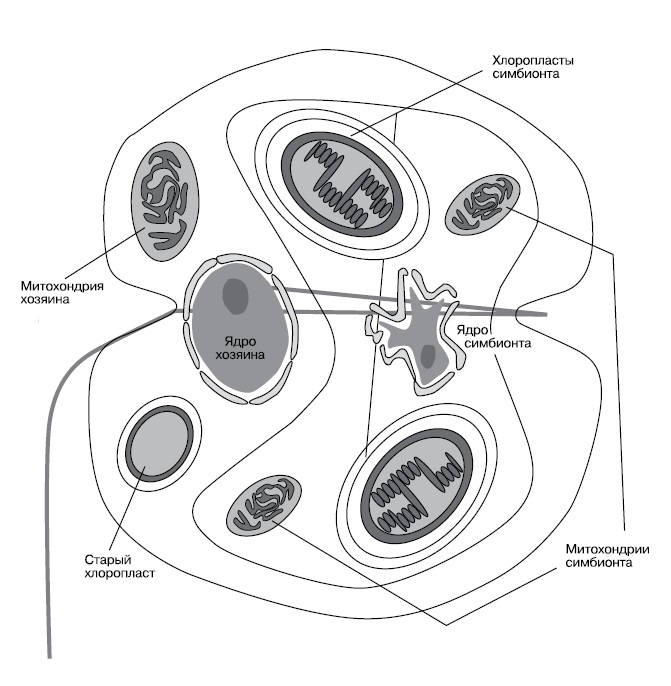
|
| 4. Схема строения клетки Kryptoperidinium (http://www.polypompholyx.com/2013/05/organism-of-the-week-14/) |
С другими причудливыми примерами перемещения хлоропластов между клетками можно ознакомиться в статье Патрика Килинга («American Journal of Botany», 2004, 91, 10, 1481—1493, doi: 10.3732/ajb.91.10.1481). Популярный рассказ о том же размещен на известном читателю «Химии и жизни» ресурсе fieldofscience.com.
И конечно, нужно заключение, раз уж мы подобрались к концу цикла. Итак, мы последовательно рассмотрели этапы биогенеза, от синтеза химических элементов в звездах и до образования эукариотной клетки. Мы видели, как сложные системы, казалось бы, неработоспособные без любой детали, могли эволюционировать из более простых благодаря смене функций. Были найдены удовлетворительные сценарии для большинства маловероятных событий на пути возникновения жизни — от появления хиральной чистоты первых биомолекул до возникновения сложно организованной клетки эукариот, объединившей в себе детали бактерий, архей и вирусов.
Мы часто повторяли, что в той или иной области наши знания не полны, поэтому свежие интересные новости на эту тему еще будут. Однако возникновение живого из неживого уже не может претендовать на звание неразрешимой загадки.
Элементы жизни: почему не кремний и не фтор
Фотохимия и «черные курильщики»
Пути восстановления углекислого газа
Происхождение белкового синтеза и генетического кода
Мир вирусов, последний общий предок и происхождение ДНК
Происхождение мембран и мембранной биоэнергетики
Закат «цинкового мира», прокариотная биосфера и происхождение фотосинтеза
Роль вирусов в происхождении клеточного ядра